







تجربههای دینی و معنوی ، همچون مراقبه، نیایش، و مناسک، در نوشتجات زیستپزشکی، روانشناختی، انسانشناختی و دینی مورد توصیف قرار گرفتهاند. تاریخ برخی از این توضیحات خاص و متون دینی به چند هزار سال پیش بازمیگردد. بهتازگی رشد فزایندهای در تعداد پژوهشهایی رخ داده است که همبستههای نوروفیزیولوژیکی و فیزیولوژیکی چنین تجربههایی را میکاوند. از چشماندازی تکاملی، چهبسا که چنین تجربههایی با رشد ساختارهای گوناگون مغز نخستیهای اولیه، و سرانجام در هوموسِیپینس، امکانپذیر شده باشند…
.
عصبروانشناسی (نوروسایکولوژی) تجربه دينی و معنوی
اندرو بی. نیوبرگ
استفانی کی. نیوبرگ[1]
ترجمه محسن زندی (پژوهشگر روانشناسی دین)
تجربههای دینی و معنوی ، همچون مراقبه، نیایش، و مناسک، در نوشتجات زیستپزشکی[2]، روانشناختی، انسانشناختی و دینی مورد توصیف قرار گرفتهاند. تاریخ برخی از این توضیحات خاص و متون دینی به چند هزار سال پیش بازمیگردد. بهتازگی رشد فزایندهای در تعداد پژوهشهایی رخ داده است که همبستههای نوروفیزیولوژیکی[3] و فیزیولوژیکی چنین تجربههایی را میکاوند. از چشماندازی تکاملی، چهبسا که چنین تجربههایی با رشد ساختارهای گوناگون مغز نخستیهای اولیه، و سرانجام در هوموسِیپینس،[4] امکانپذیر شده باشند. گمان میرود که پیدایش سازوکارهای مغزی «دینزا»[5] در هوموسِیپینس، از لحاظ تاریخی با رشد ناگهانی سنتهای دینی همزمان شده باشد، که پیوسته به راهیابی در جوامع انسانی از زمانهای پیشاتاریخ ادامه دادهاند. در پرتو این الگوی تکاملی، همبستههای نوروبیولوژیکی [عصبزیستشناختی] و نوروسایکولوژیکی[6] [عصبروانشناختی] تجربههای دینی و معنوی کمکم شناسایی شدهاند. همچنین با توجه به دیگر پژوهشهای مرتبط در نوروبیولوژی،[7] الگوی پیچیدهتری از رخدادهای نوروسایکولوژیکی در طی تجربههای دینی و معنوی را میتوان به دست آورد. به بیان دقیقتر، کارکرد مغز را میتوان با توجه به اتصالات داخلیاش با سایر قسمتهای بدن که متأثر از دستگاه عصبی خودمختار و همچنین دستگاه غدد درونریز عصبی هستند، مورد بررسی قرار داد. بررسی ارتباط بین فرآیندهای شناختی در مغز و دستگاه عصبی خودمختار، چهبسا به فهمی همهگیرتر از گونههای فراوان تجربههای معنوی بینجامد، که دامنه آنها در گسترهای از «حیرت» تا حالات عرفانی شدید کشیده میشود. در نتیجه، از آثار کنونی میتوان به عنوان بنیانی برای پدیدآری الگویی نوروسایکولوژیکی برای راهبری پژوهشهای آتی پیرامون نوروبیولوژی تجربههای دینی و معنوی استفاده کرد. پیشرفتهترین روشهای تصویربرداری مغزی که میتوانند شبکۀ انتقالدهندههای عصبی گوناگون را بررسی کنند، و نیز دیگر سنجههای فیزیولوژیکی، را میتوان در تحقیق بر کارکرد مغز در طی تجربههایی چون مراقبه، دعا و نیایش، و تجربههای آیینی به کار گرفت.
این فصل به واکاوی نوروسایکولوژی تجربههای دینی و معنوی میپردازد و دربرگیرنده وارسی کوتاهی از جنبههای پدیدارشناختی چنین تجربههایی، و نیز ترکیب دادههای موجود برای پدیدآری الگویی فراگیر است که بتواند شالودهای برای تحلیلهای آتی در ریشههای بیولوژیکی این تجربهها، و ارتباط بین این تجربهها و بهباشی روانشناختی بهدست دهد.
تکامل مغز و تجربه معنوی
فرایند تکامل به پیدایش پیوندهای پیچیده عصبی انجامیده است که در نیمکُرههای مغزی وجود دارند. همچنین مراکز عالیتر در مغز با ساختارهای ابتداییتری چون دستگاه لیمبیک [کناری] در پیوندند. رویهم رفته، مغز در فرایند تکامل پیچیدگیاش را تکامل داد تا تواناییهای پیشرفتهای را برای بازنمود نظم محیط بیرونی و حل مشکلات شناختیِ لازم برای بقا را در اختیار انسانها بگذارد. افزون بر جنبههای صرفاً شناختی، تکامل مغز به اجتماعیشدن انسان انجامید. این توانایی برای پدیدآری واحدهای خانواده، گروه، اجتماعات و جوامع، امتیاز شگرف تکاملیای را دربرداشت. از اینرو، پرسش ما در اینجا این است که چگونه این تغیرات تکاملی در مغز، به پیدایش و رشد تجربههای معنوی، دین، مناسک و آیینها منجر میشود؟
بهلحاظ کارکردی مغز را از میتوان به چند کارکرد بنیادینِ شناختی تقسیم نمود (d’Aquili, 1978, 1983, 1986). ما پیشتر این کارکردها را “عملگرهای شناختی”[8] نامیدهایم. واژه عملگر شناختی، تنها به سازوکارهای نوروفیزیولوژیکیای اشاره دارد که زیربنای برخی زمینههای گسترده کارکرد شناختی را تشکیل میدهند. از اینرو، این عملگرها به معنای دقیق کلمه وجود ندارند؛ بلکه، (فرض آنها) برای بررسی کارکرد کلی مغز سودمند است. مفهوم عملگرهای شناختی، شبیه به مفهوم پراستفادهتر بخشها[9] یا واحدهای شناختی است. با اینهمه، بنابه فرض عملگرهای شناختی با کارکردهای کلیتر مغز اشاره دارد. این عملگرهای شناختی شامل انتزاع کلی [یا کل] از جزئیات، ادراک علیت در حقیقت خارجی، ادراک پیوستگی فضایی یا زمانی در حقیقت خارجی، و مرتبسازی پارههای واقعیت در زنجیره علّی است. این کارکرد اخیر، احتمالاً همان است که الگوهای تبیینی[10] از جهان خارج را ایجاد میکند؛ حال، خواه تبیین علمی باشد و خواه رازآلود و عرفانی. در اینجا مجال توضیح جزئیات زیرلایههای فیزیولوژیکی و شبکههای نوروآناتومیکی[11] همه این عملگرها نیست. با اینهمه، توضیحی مختصر از چند عملگر میتواند در تشریح نوروسایکولوژی تجربههای دینی و معنوی سودمند باشد.
عملگر علّی دستاندرکار ترتیبدهیِ علّی عناصر واقعیت است که از ادراکات حسی انتزاع میشوند (d’Aquili, 1978). عملگر علّی کارکردش را از لوب آهیانهای تحتانی در نیمکره چپ، برجستگی پیشین لوبهای پیشانی، بیشتر در نیمکره چپ، و از پیوندهای داخلی دوسویه عصبی میگیرد(Luria, 1966; Pribram, 1973). گمان میرود که عملگر علّی اهمیت بنیادینی در پدیدآری تجربهها و مفاهیم دینی و معنوی داشته باشد (d’Aquili, 1978). این عملگر هر پاره مفروض از واقعیت را در یک پیوستار علّی ذهنی سامان میدهد و آن را تا ایستگاهِ آغازینش ردّگیری میکند. با تأمل در ویژگی (به گمان) فراگیر انسان در قرار دادنِ علت برای هر تکه از واقعیت، میتوان گفت که حتی اگر ایستگاه آغازین سلسلۀ علی یادشده توسط دادههای حسی به دست نیاید، عملگر علی بهطور خودکار آن ایستگاه آغازین علّی را میزایاند (d’Aquili & Newberg, 1993). علم غربی، مسلّم انگاشتن ایستگاهی آغازین یا علّت نخستین را برای تکههای واقعیت نمیپذیرد، مگر اینکه آن علت نخستین یا مورد مشاهده قرار گیرد، و یا اینکه بتوان بدون واسطه آن را از مشاهده [حسی] استنتاج نمود. در زندگی روزمرّه (غیرعلمی)، عملگر علّی بهسادگی یک ایستگاه نخستین یا علّت نخستین برای هر تکه از واقعیت میسازد. به گمان ما، هنگامی که هیچ تبیین علّی علمی یا مشاهدهای برای تکهای از واقعیت به دست نمیآید، خدایان، نیروها[ی برین]، ارواح یا برخی دیگر از ساختارهای سبَبی بهطور خودکار توسط عملگر علّی پدید میآیند[12](d’Aquili & Newberg, 1997). اگر اینگونه باشد، وقتی که مشاهدات ما ایستگاه آغازین را نشان ندهد، چهبسا عملگر علّی بهطور خودانگیخته وارد میشود و پایانه علّی نخستینی را برای واقعیت بیرونی میزایاند.
اگر این سخن درست باشد که عملگر علّی واقعیت را لزوماً تحلیل میکند، در آنصورت، انسانها چارهای جز ساختن اسطورههایی آکنده از نیرویی شخصوار برای تبیین جهان پیرامونشان ندارند. اسطورهها چهبسا ماهیتی اجتماعی داشته باشند، و یا برحسب خوابها، خیالبافیها، یا دیگر جنبههای خیالآلود شخص، پدیدهای با رنگوبوی فردی باشند. با اینوجود، انسانها تا آنجا که به امکان وجودشان در رویاری با جهان ناپایدار و اعتمادناپذیر پیرامونشان آگاهی دارند، اسطورههایی برای شناسایی راه و جایگاه خود در جهان هستی و سازگاری با زیستگاه پیرامونشان پدید میآورند. از اینرو، خدایان، ارواح، دیوها و فرشتهها، یا دیگر سرچشمههای قدرت شخصواری را میآفرینند که میتوانند با آنها قرارداد بسته و داد و ستد نمایند، تا در نتیجه، مهار زیستبومِ ناپایدار و اعتمادناپذیر خود را بهدست گیرند.
دومین عملگری که به گمان ما اهمیت ویژهای در تجربه معنوی دارد، عملگر کلنگر[13] است. عملگر کلنگرِ پیشنهادی به انسان اجازه میدهد که واقعیت در قالب یک کُل یا گشتالت[14] بنگرد و انتزاع یک چهارچوب بافتاریِ گستردهتر[15] را از جزءها یا افراد میسر میسازد. عملگر علی، بهگمان بسیار، در لوب آهیانهای در نیمکره راست (یا نیمکره ناچیره)،[16] و به سخن دقیقتر، در قطعه آهیانهای بالاتر خلفی و ناحیههای مجاور جای دارد، که امروزه میدانیم در پدیدآری فهم گشتالتی پیرامون دادههای حسی و مفاهیم انتزاعی گوناگون دخالت دارند (Bogen, 1969; Gazzaniga & Hillyard, 1971; Nebes & Sperry, 1971; Sperry, Gazzaniga, & Bogen, 1969). توجه به این نکته نیز جالب است که این ناحیه در مقابل ناحیهای در نیمکره چپ قرار دارد که زیرلایه نوروآناتومیکی[17] را برای عملکردهای منطقیـدستوری فراهم میکند. در نتیجه، لوب آهیانهای راست در رویکرد کلنگرانه به چیزها، و لوب آهیانهای چپ در فرایندهای فروکاهشگرایانهتر[18] پادرمیانی دارند. ما در زیر به بررسی این مسأله میپردازیم که این ساختارهای گوناگون، و ساختارهای مرتبط، چگونه میتوانند بهطور خاصتر در تجربههای دینی و معنوی دخالت داشته باشند.
روشهای دستیابی به تجربههای معنوی
در بررسی الگوی نوروسایکولوژیکی تجربههای دینی و معنوی، واشکافی اینکه چگونه چنین تجربههایی بهدست میآیند، بسیار مهم است. به گمان ما، دو مقوله کلی در روشهای دستیابی به چنین تجربههایی وجود دارد: مناسک گروهی،[19] و تأمل فردی،[20] از قبیل نیایش یا مراقبه. تحلیل پدیدارشناختی این دو نوع عمل نشان میدهد که آنها جنسی مشترک (اگرچه، نه در شدت و حدت) دارند: 1) تخلیههای هیجانی ادواری که احساسِ ذهنی حیرت، صلح، آرامش، یا خلسه (از خود بیخودی) را در خود دارند؛ و 2) درجات متغیر تجربه وحدت، که با تخلیههای هیجانی یادشده متناظر است (d’Aquili & Newberg, 1993). این تجربههای وحدتگرا متشکل از کاهش احساس یا آگاهی از مرز میان «خود» و جهان بیرونی است (d’Aquili, 1986; d’Aquili & Newberg, 1993; Smart, 1958, 1967, 1969; Stace, 1961). بُعد اخیر همچنین میتواند به احساس وحدت میان دیگر افراد واقع در دایرۀ ادراک بینجامد، و بهموجب آن احساس باهَمِستانی[21] پدید آید. سرانجام، تجربههای وحدت میتواند به برچیدن همه مرزهای وجودِ ناپیوسته و گسسته منتهی شود، که در نتیجه، حالت یگانگی نامتمایز (یا آنچه را که ما وجود واحد محض[22] نامیدهایم) را به بار آوَرَد (AUB; d’Aquili & Newberg, 1999). باید توجه داشت که تجربههای آیینی گروهی و مراقبه فردی تا حدی همپوشانی دارند؛ تا آنجا هر یک میتواند نقشی را در پیدایش دیگری بازی کند. در واقع، چهبسا که مناسک آیینی شخص “میانهای”[23] را پدید آورَد که به تجربه رازآلود و عرفانی دست مییابد («میانه»، در تمایز با آنانی است که بهطور منظم و فراوان تأملات شدید دارند؛ از قبیل راهبان و زاهدان تارک دنیا و بهشدت متدین). این سخن بههیچ روی بدان معنا نیست که تجربه رازآلود و متأملانه در برابر تأثیرات مناسک آیینی بیتفاوت است. دقیقاً بهدلیلِ تجربههای وحدت شدیدِ ناشی از مراقبه است که تأثیرپذیری عارفان از مناسک آیینی در عمل ممکن است بیشتر از شخص میانه باشد؛ اگرچه این مطلب هنوز اثبات نشده است. چه بسا بتوان نتیجه گرفت که مناسک آیینی، در بیشترین اثربخشیاش، شگردی بیاندازه نیرومند (حال چه خوب، چه بد) است. همچنین، به دلیل ابعاد ذاتاً عمومی آن، معمولاً اهمیت اجتماعی بسیار بیشتری نسبت به مراقبه یا تأمل فردی دارد. اگرچه مراقبه و تأمل میتواند حالات وحدت شدیدتر و گستردهتری را ایجاد کنند (که به برقهای نسبتاً کوتاه تولیدشده توسط آیین گروهی تشبیه شدهاند)، اما کمابیش هماره تجربههایی انزواطلب و عزلتگزین هستند.
بهنظر میرسد که مناسک آیینی از نظر اخلاقی فنونی خنثی هستند؛ بدان معنا که هم میتوانند در اهداف سازنده و مثبت و هم منفی و ویرانگر مورد استفاده قرار گیرند. از اینرو، آیین با تکیه بر اسطورهای که هم دربردارنده و هم بیانگر مناسک آیینی است، میتواند جنبههای ساختاری یک اجتماع[24] را گسترش دهد یا بکاهد، و رفتار پرخاشگرانه را برانگیزاند و یا بکاهاند. با بهکارگیری نظریه ترنر (1969) درباره باهَمِستانی (بهعنوان تجربه نیرومند اجتماعیـاتحادی که بیشتر در نتیجه مناسک آیینی رخ میدهد)، میتوانیم بگوییم که اگر یک اسطوره تجسّمش را در آیینی به دست آورَد که تجربه وحدت را برای گروه یا قبیلهای خاص معین سازد، در آن صورت، نتیجه فقط یگانگی و باهمِستانی قبیلگی[25] است. این سخنِ درستی است که با وحدتبخشی به تجربه پدیدآمده توسط آیین، پرخاشگری و تجاوز در میان گروه به حداقل و یا صفر میرسد. با اینهمه، حتی اگر تجاوز و پرخاشگری درونگروهی کاسته شود، چهبسا که این تجربۀ وحدت درونگروهی «تنها» به تأکید بر یکپارگی گروه در تمایز و تقابل با دیگر گروهها کمک کند، و در نتیجه، به افزایش پرخاشگریهای میانگروهی بینجامد. البته، گاه گستره اسطوره و تجسمبخشی آیینی به آن شامل همه اعضای وابسته به یک دین، ملتـکشور، یک ایدئولوژی، همه انسانها، و یا همه واقعیتها میشود. روشن است به هر اندازه که گستره آنچه در تجربه اتحادی منظور میشود بیشتر باشد، رفتار پرخاشگرانه نیز کاستی میگیرد. در واقع، اگر این سخن درستی باشد که مناسک آیینی گاه میتواند بر تنِ اسطوره وحدت همه موجودات گوشت برویاند، در آن صورت فردی که چنین آیینی را انجام میدهد چهبسا که احساس زودگذر وحدت همۀ اشخاص را تجربه کند. چنین تجربه اسطورهـآیینی[26] چیزی شبیه به حالات مراقبهای، مانند آگاهی کیهانی بوک (1961) یا حتی ،AUB[27] است. با اینهمه، متأسفانه در تجربههای قومنگاشتی چنین گستره برجستهای از وحدت در مناسک گروهی بسیار نادر بوده است.
الگویی نوروسایکولوژیکی از تجربههای دینی و معنوی
الگوی توصیفشده در زیر، الگوی مفصلتری از الگوی پیشتر توصیفشدهای است، که اکنون مطالعات تازهای از تصویربردازی مغزی، شیمی اعصاب،[28] هورمونی، و فیزیولوژیکی را نیز در خود دارد (Newberg & Iversen, 2003). هدف این الگو بنای شالودهای است که از راه آن بتوان گونههای فراوانِ تجربهها و اعمال دینی را بررسی و مقایسه نمود. همانگونه که نمودار 11.1 نشان میدهد، این الگو با کُرتکس پیشپیشانی شروع میکند، و شماری از برهمکنشهای پیچیده با تالاموس، لوب آهیانهای بالاتر پشتی،[29] دستگاه لیمبیک، و دستگاه عصبی خودکار را بیان میکند. همچنین، شماری از انتقالدهندههای عصبی[30] تحریکی و بازداشتی را میتوان نشان داد که در چنین اعمال و تجربههایی نقش بازی میکنند. دوپامین، سروتونین، اَسِتیل کولین، و چند مولکول دیگر میتوانند با جنبههای گوناگون پدیدارشناختی چنین تجربههایی در پیوند باشند؛ همه اینها در این الگو واکاوی میشوند. چهبسا که سازوکارهای خاص، بر اساس عمل، آیین، سنت، و یا فردی خاص تا اندازهای متفاوت باشند. با اینهمه، این الگو با تمرکز بر پدیدارشناسی چنین تجربههایی، اطلاعاتی درباره گوناگونی تجربهها، خواه حسی، شناختی یا عاطفی، در اختیار مینهد که میتوانند با تجربههای دینی و معنوی همپیوند باشد. از آنجا که میزان کمابیش گستردهای از دادهها در دسترس بودند، این الگو عمدتاً با به کارگیری دادههای بهدستآمده از پژوهشهایی ایجاد شد که بیشتر بر اعمال مراقبهای تمرکز دارند. این الگو را چهبسا بتوان برای گونههای بسیار فراوان اعمال و تجربهها بهکار گرفت.
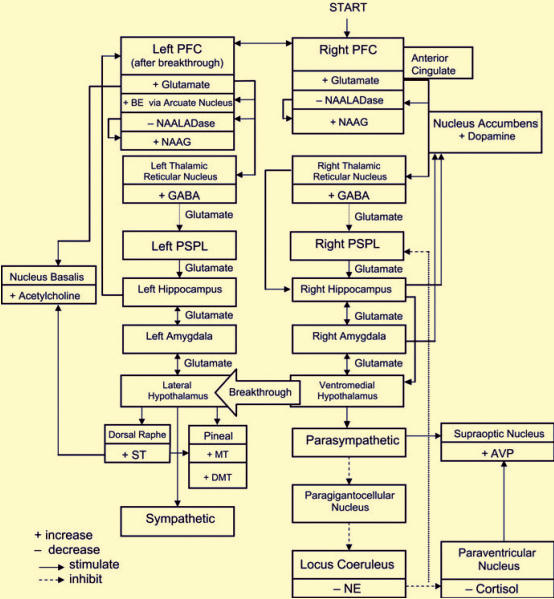
برانگیختگی کرتکس پیشپیشانی و کمربندی[31]
بیشتر اعمال مراقبهای، دعا یا دیگر اعمال متأملانه، مستلزم حدی از توجه مستمر و بیوقفه است. این حفظِ مستمر توجه میتواند با تمرکز بر تصور یک ابژه، منتره [سرودههایی در آیین هندو]، نیایش، یا برخی کانونهای معنوی دیگر انجام شود. پژوهشهای مبتنی بر تصویربرداری مغزی نشانگر آنند که فعالیتها و تکالیف ارادی و حسابشده که نیازمند حفظ بیوقفۀ توجه هستند، با فعالیت در کرتکس پیشپیشانی[32] (PFC)، بهویژه در نیمکره راست شروع میشود (Frith, Friston, Liddle, & Frackowiak, 1991; Ingvar, 1994; Pardo, Fox, & Raichle, 1991; Posner & Petersen, 1990). شکنج کمربندی [سینگیولِیت][33] نیز نشان داده شده است که در تمرکز توجه، احتمالا با PFC، همکاری دارد( Vogt, Finch, & Olson, 1992). در نتیجه، از آنجایی که اعمال معنوی نیازمند تمرکز شدید توجه هستند، این فرضیه درست بهنظر میآید که الگوی مورد نظر برای مراقبه، با برانگیختگی کرتکس پیشپیشانی (بهویژه در نیمکره راست)، و نیز شکنج کمربندی شروع کند. این ایده، با فعالیت فزاینده مشاهدهشده در این بخشها، در چندین مطالعه تصویربرداری مغزی بر روی گونههایی از انواع ارادی مراقبه، ، حمایت شد؛ از جمله مطالعاتی که در آزمایشگاه خودِ ما انجام شد و در آن هشت مراقبهکننده بودایی تبتی در خط پایه و در حال مراقبه مورد بررسی قرار گرفتند (Newberg et., al 2001 ). تحلیل کمّی، فعالیت فزایندهای را در کرتکس پیشپیشانی بهطور دوسویه ( اگرچه بیشتر در نیمکره راست) و شکنج کمربندی در هنگام مراقبه نشان داد. از اینرو بهنظر میرسد که مراقبه با برانگیختگی کرتکس پیشپیشانی و کمربندی شروع میشود، که با ارادۀ وضوحبخشی به افکار ذهنی و یا تمرکز بر یک موضوع درپیوند هستند. با اینهمه، یک پژوهش مبتنی بر پرتونگاری با نشر پوزیترون[34] (PET) پیرامون گونه هدایتشدهای از مراقبه، افزایش فعالیت پیشپیشانی را نشان نداد؛ هرچند، پژوهش تازهای در مقایسه با تولید کلام درونی یا ارادی، کاهش فعالیت قدامی را در طی تولید کلامِ هدایتشده از بیرون نشان داد (Crosson et al., 2001). در نتیجه، برانگیختگی پیشپیشانی و کمربندی میتواند با جنبههای ارادی مراقبه درپیوند باشد.
برانگیختگی تالاموس، بهعنوان بخشی از شبکه توجهی
تالاموس یک ایستگاه تقویتی مهم در مغز است که دیگر ساختارها را پیوند داده، و نیز پردازش مرتبه بالا را به بخشهایی از مغز انتقال میدهد که به کارِ هیجان میآیند؛ و دستآخر، تالاموس فرایندهای گوناگون فیزیولوژیکی را سامان میبخشد. چند پژوهش بهروی حیوانات نشان داده است که کرتکس پیشپیشانی بههنگام برانگیختگی، هسته مشبک تالاموس را عصبگیری[35] میکند ( Phillipson, 1988&Cornwall)؛ بهویژه بهعنوان بخشی از یک شبکه توجهی یکپارچهتر (Portas et al., 1998). چنین برانگختگیای چهبسا با تولید و توزیع انتقالدهنده عصبی تحریکی گلوتامات توسط کرتکس پیشپیشانی انجام شود، که نورونهای کرتکس پیشپیشانی آن را برای انتقال و رابطه میان خودشان، و عصبگیری کردن دیگر ساختارها مغزی بهکار میبرند (Cheramy, Romo & Glowinski, 1987 ). تالاموس، خودش جریان دادههای حسی بهسوی بخشهای پردازش قشری را از طریق برهمکنشهایش با هستههای زانویی جانبی و خلفی جانبی[36] اداره میکند و گمان میرود که از شبکه گلوتامات نیز برای فعال کردن نورونها در دیگر ساختارها استفاده میکند (Armony & LeDoux, 2000). میدانیم که هسته زانویی جانبی دادههای خام بصری را از جِهاز بینایی دریافت میکند و آن را از مسیری معین به کرتکس مخطط[37] برای پردازش میفرستد. هسته خلفی جانبی تالاموس، دادههای حسی را برای قطعه آهیانهای فوقانی خلفی[38] (PSPL) فراهم میکند، که برای موقعیتیابی فضایی بدانها نیاز دارد (Bucci, Conley, & Gallagher, 1999).
هسته مشبک بههنگام برانگیختگی، انتقالدهنده عصبی بازدارنده گاما آمینوبوتریک اسید (GABA) به سوی هستههای خلفی جانبی و زانویی ترشح میکند، که درونداد به PSPL و کانونهای بصری را متناسبِ با برانگیختگی هسته مشبک قطع میکند (Destexhe Contreras, & Steriade, 1998). در طی مراقبه، بهخاطر افزایش فعالیت درPFC، بهویژه در نیمکره راست، بهلحاظ نظری باید افزایش توأمانی در فعالیت هسته مشبک تالاموس انجام پذیرد. درحالیکه مطالعات تصویربرداری مغزی از مراقبه راهحلی برای تمایز هستههای مشبک نداشتهاند، اما تک مطالعه تازه توموگرافی محاسبهشده نشر فوتون[39](SPECT) توسط ما، افزایش کلی فعالیت تالاموسی را نشان داد که متناسب با سطوح فعالیت درPFC بود. این امر با برهمکنش ویژه میان PFC و هستههای مشبک سازگار است (اما تأیید نمیکند). اگر برانگیختگی PFC نیمکره راست، علت افزاش فعالیت در هسته مشبک در طی مراقبه باشد، در آنصورت چهبسا نتیجه کاهشِ درونداد حسیای باشد که به PSPL وارد میشود. چندین مطالعه افزایش سرم GABA [گابا] را در هنگام مراقبه نشان دادهاند، که گمان میرود افزایش فعالیت GABA مرکزی را بازتاب میدهد (Elias, Guich, & Wilson, 2000). این آورانبرداری[40] کارکردی همپیوند با افزایشِ GABA بدین معناست که محرک حواسپرتکننده و مزاحم بیرونی کمتری به کرتکس بینایی و PSPL میسد،؛ که این پدیده با افزایش حس تمرکز در طی مراقبه رخ میدهد.
همچنین باید این نظریه را یادآوری نمود که شبکه دوپامینی[41] از طریق هستههای قاعدهای[42] در تنظیم شبکه گلوتاماتی[43]و برهمکنش میان کرتکس پیشپیشانی و ساختارهای زیر قشری شرکت میکند. یک پژوهش تازه مبتنی بر PET [پرتونگاری با نشر پوزیترون] آهنگ دوپامین را در طی مراقبه نیدرَه یوگا[44] مورد اندازهگیری قرار داد و افزایش معنیدار سطوح دوپامین را در طی عمل مراقبه نشان داد (Kjaer et al., 2002). کایر و همکارانش این فرض را مطرح کردند که این افزایش چهبسا با قطع کنش و واکنشهای قشری و زیرقشری همپیوند است که به کاهش کلی آمادگی برای کنش میانجامد؛ که این نیز همپیوند با اینگونه خاص از مراقبه است. در مطالعات آینده لازم است که نقش دوپامین در طی اعمال مراقبهای، و نیز برهمکنشهای بین دیگر شبکههای انتقالدهنده عصبی به دقت بررسی و روشن شود.
آوران برداریPSPL
پژوهشها نشان دادهاند که PSPL در تحلیل و یکپارچهسازی دادههای بصری، شنیداری، و زیباییشناختی جسمی مرتبه بالا دخالت دارد ( Adair, Gilmore, Fennell, Gold & Heilman, 1995; see also Joseph, 1990). همانگونه که پیشتر نیز گفتیم، گمان میرود که این ساختار نقش بنیادینی در فرآیندهای شناختی کلنگرانه و نیز تقلیلگرایانه بازی میکند. PSPL همچنین در شبکه توجهی پیچیدهای که دربرگیرنده PFC و تالاموس است، شرکت دارد (Fernandez-Duque & Posner, 2001 ). PSPL با دریافت درونداد شنیداری و دیداری از تالاموس، قدرت مییابد که به ایجاد تصویر سهبُعدی بدن در فضا، و ایجاد احساس مختصات فضایی که بدن در جهت آن قرار میگیرد، و تمییز میان چیزها کمک کرده و در خصوص ابژههایی که میتوانند مستقیماً بهچنگ آیند و مورد استفاده و دستکاری قرار گیرند تأثیر بگذارد (Mountcastle, Motter, & Anderson, 1980; Lynch, 1980). چهبسا این کارکردهای PSPL برای تمایز نهادن میان خود و جهان بیرون، مهم است. این را نیز باید گوشزد کنیم که پژوهشی تازه بر آن است که لوب گیجگاهی فوقانی چهبسا نقش بنیادینیتری در بازنمایی فضایی بدن بازی میکند؛ اگرچه این دستآورد با گزارشهای دیگر ثابت نشده است (Karnath, Ferber, & Himmelbach, 2001). به هر روی همچنان باید به جستجوی پاسخی برای این بود که چه پیوند حقیقیای میان لوبهای آهیانهای و گیجگاهی از نظر بازنمایی فضایی وجود دارد؟
بههر روی، به گمان ما آورانبرداری از این بخشهای جهتیاب مغز، مفهوم مهمی در فیزیولوژی مراقبه است. برای نمونه، اگر آورانبرداری PSPL با تأثیرات شبکۀ اسید گاما آمینوبوتریک[45] هسته مشبک رخ دهد، ممکن است که شخص شروع به از دست دادن توانمندی معمول خود در بازشناسی فضایی خود کند. این ایده با یافتههای بالینی در بیماران مبتلا به سندروم بالینت[46] تأیید میشود، که در آن آسیب لوب آهیانهای موجب دشواری چشمگیر مبتلایان در بازشناسی خودشان در فضای سهبعدی میشود. البته، چهبسا که تأثیرات مراقبه گزینشیتر باشد و «احساسِ از خود» را ویران نسازد؛ بلکه ادراک[47] آن را دگرگون سازد. همچنین، آورانبرداری PSPL با دو پژوهش مبتنی بر تصویربرداری که کاهش فعالیت در این بخش را در طی مراقبه شدید نشان میدهند، مورد تأیید قرار گرفته است (Newberg et al., 2001; Herzog et al., 1990–1991). همچنین مطالعه SPECT صورتگرفته توسط ما، همبستگی میان افزایش فعالیت در تالاموس و کاهش فعالیت در PSPL را نشان داد. پیامد این امر آن است که هرچه فعالیت PFC فرد افزایش یابد، آورانبرداری PSPL بیشتر میشود. از اینرو، چهبسا محققی این فرض برایش ایجاد شود که تمرکز هرچه شدیدتر و عمیقتر باشد، احتمال اینکه شخص سرانجام به حالتهای وحدتگون دست یابد بیشتر است.
برانگیختی هیپوکامپ و آمیگدال در طی اعمال معنوی
افزون بر فعالیت پیچیده قشریـتالاموسی، میتوان انتظار داشت که اعمال مراقبهای و معنوی فعالیت دستگاه لیمبیک را نیز تغییر میدهند؛ بهویژه از هنگام تحریک ساختارهای لیمبیکیِ مرتبط با تجربههای مشابه با آنچه در طی این اعمال توصیف میشوند (Fish, Gloor, Quesney, & Olivier, 1993; Saver & Rabin, 1997). هیپوکامپ دست بهکار میشود تا انگیختگی و پاسخدهی قشری را از راه پیوندهای داخلی پهناور و توانگر با کرتکس پیشپیشانی، بخشهای نئوکورتکس،[48] آمیگدال [بادامه] و هیپوتالاموس سامانبخشی و تعدیل نماید (Joseph, 1990). معلوم شده است که تحریک هیپوکامپ انگیختگی و پاسخدهی قشری را میکاهد؛ البته، اگر انگیختگی قشری ابتدا در سطح پایینی باشد، در آنصورت تحریک هیپوکامپ معمولاً فعالیت قشری را میافزاید (Redding, 1967). توانایی هیپوکامپ در برانگیزی یا بازداری فعالیت یاختههای عصبی در ساختارهای دیگر چهبسا بهترتیب مبتنی بر شبکههای گلوتامات و GABA باشد. (Armony & LeDoux, 2000). ایدۀ ما در الگوی نوروسایکولوژیکیمان درباب مراقبه آن است که در طی مراقبه یک آورانبردای جزئی از PSPL نیمکره راست وجود دارد. این آورانبرداری همچنین بهخاطر تنظیم وارونه هیپوکامپ نسبت به فعالیت قشری، میتواند به تحریک هیپوکامپ راست بینجامد. همچنین، اگر تحریک مستقیم و همزمان هیپوکامپ راست از طریق تالاموس (بهعنوان بخشی از شبکه توجهی شناختهشده) باشد، و از گلوتامات متأثر شود، بهنظر ما در آنصورت توانگیری قدرتمندی از تحریک هیپوکامپ راست رخ میدهد. چهبسا فعالیت هیپوکامپی راست سرانجام کارکرد تحریککنندگی PFC بر تالاموس را از طریق هسته اکامبنس[49] افزایش دهد، که آماده قطع دادههای عصبی از PFC به تالاموس از راه تأثیرات تعدیلکنندگی عصبی دوپامین است ( Newman & Grace 1999).
هیپوکامپ تأثیر بسیاری بر آمیگدال مینهد، بهطوریکه آنها همدیگر را کامل میکنند و در ایجاد توجه، هیجان و گونههای خاصی از تصویرسازی ذهنی با یکدیگر برهمکنش دارند (Joseph, 1990). بهنظر میرسد که بسیاری از سامانبخشیهای پیشپیشانی هیجان از طریق هیپوکامپ و اتصالاتش با آمیگدال انجام میپذیرد (Poletti & Sujatanond, 1980). بهخاطر این برهمکنش دوسویه میان آمیگدال و هیپوکامپ، به گمان ما برانگیختگی هیپوکامپ راست در طی مراقبه، احتمالاً آمیگدال جانبی را نیز تحریک میکند. نتایج تصویربرداری با تشدید مغناطیسی کارکردی (fMRI)[50]، توسط لازر و همکارانش (2000) باور به افزایش فعالیت در بخشهایی از آمیگدال و هیپوکامپ در طی مراقبه را مورد تأیید قرار میدهد.
تغییرات هیپوتالاموس و دستگاه عصبی خودکار
میدانیم که هیپوتالاموس بهطور گستردهای با دستگاه لیمبیک در پیوند است. پژوهشها نشان دادهاند که تحریک آمیگدال جانبی راست منجر به تحریک قسمت [هسته] وانترومدیان ِهیپوتالاموس با تحریک بعدیِ دستگاه پاراسمپاتیک پیرامونی میشود (Davis, 1992). افزایش فعالیت پاراسمپاتیک با نخستین احساس ذهنی آرمیدگی [ریلکسِیشِن]، و سرانجام، با احساس سکون و آرامش عمیقتر، در پیوند است. همچنین، برانگیختگی دستگاه پاراسمپاتیک علت کاهش ضربان قلب و ضربآهنگ تنفسی است. همه این واکنشهای فیزیولوژیکی در هنگام مراقبه مشاهده شدهاند (Jevning, Wallace, & Beidebach, 1992).
برای نمونه، وقتی ضربآهنگ تنفس و ضربان قلب کاهش مییابد، هسته پاراگیگانتوسِلیولرِ مدّولا[51] از کار دست میکشد تا لوکوس سرولوس[52] (LC) پل دماغی را عصبگیری کند. لوکوس سرولوس، نوراپی نفرین (NE) را تولید و توزیع میکند (Foote, 1987)؛ تعدیلکننده عصبیای که پذیرندگیِ بخشهای مغزی برای دروندادهای حسی را با تقویت محرکهای نیرومند افزایش میدهد؛ درحالی که بهطور همزمان برانگیختگیهای ضعیفتر و «پارازیت» سلولیای[53] که پایینتر از آستانه برانگیختگی باشد را بیرون نگاه میدارد (Waterhouse, Moises, & Woodward, 1998). کاهش تحریک لوکوس سرولوس منجر به کاهش سطح نوراپی نفرین میشود (Van Bockstaele & Aston-Jones, 1995). عموماً مشاهده شده است که اختلال فرآوردههای کاتکولامینهایی از قبیل نوراپی نفرین و اپی نفرین در ادرار و در پلاسما در طی مراقبه کاهش مییابد (Walton, Pugh Gelderloos, & Macrae, 1995)، که ممکن است صرفاً تغییر نظاممند توازن دستگاه عصبی خودمختار را نشان میدهد. هرچند، این امر با کاهش مُخّی[54] سطوح نوراپی نفرین هم ناسازگار نمیباشد. الگوی پیشنهادی ما نشان میدهد که در طی مراقبه، کاهش شلیک هسته پاراگیگانتوسلولر، عصبگیریاش از لوکوس سرولوس را کاهش میدهد؛ که میدانیم این نیز بهنوبه خود نوراپی نفرین را برای PSPL و هسته خلفی جانبی تهیه میکند (Foote, 1987). از اینرو، کاهش نوراپی نفرین تأثیر درونداد حسی بر PSPL را با کمک به آورانبرداری آن میکاهد.
همچنین، لوکوس سرولوس نوراپی نفرین کمتری را به هسته دور بطنی[55] هیپوتالاموس میرساند. هسته دور بطنی هیپوتالاموس هماره هورمون رهاکننده کورتیکوتروپین (CRH) را در واکنش به تحریک عصبی توسط نوراپی نفرین از لوکوس سرولوس ترشح میکند (Ziegler, Cass, & Herman, 1999). این CRH هیپوفییز قدامی را به رهاسازی هورمون محرک قشر غدههای فوق کلیه [ادرنوکوتیکوتروپیک][56] (ACTH) تحریک میکند (Livesey, Evans, Mulligan, & Donald, 2000). ACTH بهنوبه خود کرتکس فوق کلیوی را به تولید کورتیزول تحریک میکند، که یکی از هورمونهای استرس بدن است (Davies, Keyon, & Fraser, 1985). کاهش نوراپی نفرین از لوکوس سرولوس در طی مراقبه احتمالاً تولید CRH توسط هسته دور بطنی، و سرانجام سطوح کوتیزول را میکاهد. بیشتر پژوهشها دریافتهاند که سطوح کورتیزول ادرار و پلاسما در طی مراقبه کاهش مییابد (Jevning, Wilson, & Davidson, 1978; Sudsuang, Chentanez, & Veluvan, 1991 ) که این یافته از این نظریه پشتیبانی میکند که کاهش کلی ترشح کورتیزول وجود دارد. این امر همچنین نتایجی برای رابطه میان اعمال مراقبهای و کاهش استرس دارد؛ زیرا کورتیزول هماره بهعنوان هورمون اصلی استرس مورد توجه میباشد.
انتظار میرود که افت فشار خونِ همپیوند با فعالیت پاراسمپاتیک، گیرندههای فشار شریانی را در طی اعمال مراقبهای آرمیده نماید، که این نیز مدولای بطنی دُمی[57] را به کاستن بازداری اسید گاما آمینوبوتریکی[58] از هسته سوپرااُپتیک هیپوتالاموس میکشاند. در شرایط خاصی، این نبودِ بازداری میتواند هسته سوپرااُپتیک را به رهاسازی هورمونِ رگ تنگکنِ آرژنین[59] (AVP) تحریک کند که بر اثر آن شریانها تنگ میشوند و فشار خون به سطح طبیعی بازمیگردد (Renaud, 1996). همچنین نشان داده شده کهAVP به حفظ کلی عاطفه مثبت کمک میکند (Pietrowsky, Braun, Fehm, Pauschinger, & Born, 1991)، ادراک از انگیختگی و خستگی را میکاهد، و بهطور معناداری تثبیت و تقویت حافظهها و یادگیریها جدید را بهبود میبخشد (1981.,(Weingartner et al. در واقع، AVP پلاسما نشان داده شده که بهطور چشمگیری در طی مراقبه افزایش مییابد (O’Halloran et al., 1985). از اینرو افزایش در AVP میتواند به کاهش احساس ذهنی خستگی، و افزایش حس انگیختگی منجر شود. همچنین میتواند به افزایش حافظه مراقبهکننده از تجربهای که در آن است کمک کند، و چهبسا تبیینی برای این پدیدار ذهنی باشند که چرا تجربههای معنوی و مراقبهای به خاطر سپرده میشوند، بهیاد میآیند و در واژگانی پرشور توصیف میگردند.
تأثیرات PFC بر شبکههای عصبشیمیایی دیگر
هنگامی که عمل معنوی ادامه مییابد، فعالیت PFC مرتبط با اراده پایدار و مُجدانه برای تمرکز توجه نیز باید ادامه یابد. بهطور کلی، هنگامی که فعالیت PFC افزایش مییابد، سطوح روبهافزایشی از گلوتاماتِ سیناپسی آزاد را در مغز تولید میکند. افزایش گلوتامات میتواند هسته آرکوئیت هیپوتالاموس را به رها کردن اندورفین[60] بتا(BE) تحریک میکند (Kiss, Kocsis, Csaki, Gorcs, & Halasz, 1997). BE یک شِبه افیون است که بیشتر توسط هسته آرکوئیت هیپوتالاموس میانی[61] ساخته میشود و در نواحی زیرقشری مغز توزیع میگردد (Yadid, Zangen, Herzberg, Nakash, & Sagen, 2000). میدانیم که BE تنفس را میکاهد، ترس و درد را کاهش میدهد، و احساس شادی و سرخوشی پدید میآورد (Janal, Colt, Clark, & Glusman, 1984). آن تأثیراتی که از حالت مراقبه توصیف میشود، میتواند حاکی از رها شدن مقداری BE همپیوند با افزایش فعالیت PFC باشد. پژوهشها دریافتهاند که مراقبه چرخههای روزانه BE و ACTH را بر هم میزند؛ گرچه بر چرخههای روزانه کورتیزول تأثیری نمینهد (Infante et al., 1998). بههر روی، گمان میرود که BE تنها واسطه چنین تجربههایی در طی مراقبه نباشد؛ زیرا صِرفِ دریافت مواد مرتبط با مرفین تجربههای عرفانی همسنگ ایجاد نمیکنند. همچنین، پژوهشی بسیار محدود نشان داد که انسداد گیرندههای افیوندار، با نالوکسن،[62] بر این تجربه، یا الکتروانسفالوگرام [موجنگار مغزی] (EEG) همراه با مراقبه، تأثیری نگذاشت (Sim & Tsoi, 1992).
گلوتامات گیرندههای انـمتیلـدیـاسپارتیت[63] (NMDAr) را فعال میکند؛ اما گلوتامات اضافی میتواند این نورونها را با فرایند تحریک بیش از حد[64] از بین ببرد (Albin & Greenamyre, 1992) به گمان ما اگر سطح گلوتات به غلظت تحریک بیش از حد در طی حالات شدید مراقبه نزدیک شود، ممکن است که مغز تولید دیپِپدیداسِ اسیدی انـاستیلیشده آلفادار[65] (که انـاَسِتیل اسپارتیل گلوتاماتِ درونزادِ ضد NMDAr[66] را به گلوتامات تبدیل میکند) را محدود کند (Thomas, Vornov, Olkowski, Merion, & Slusher, 2000). افزایش NAAG، سلولها را از آسیب تحریک بیش از حد محافظت میکند. با اینحال، یک نتیجه فرعی مهم وجود دارد؛ NAAG بازدارنده NMDAr بهلحاظ کارکردی قابل مقایسه با توهمزاهای ناپیوندی کتامین،[67] فن سیکلیدین و اکسید نیترو[68] است (Jevtovic- Todorovic, Wozniak, Benshoff, & Olney, 2001). این متضادهای NMDAr حالتهای گوناگونی پدید میآورند که ویژگی آنها را میتوان در قالب تجربههای اسکیزوفرنیک یا تجربههای عرفانیای همچون خلع بدن،[69] و تجربههای نزدیک به مرگ دید (Vollenweider et al., 1997).
فعالیت دستگاه عصبی خودمختار[70]
در اوایل دهه 1970، گلهورن[71] و کیلی[72] الگویی از فرآیندهای فیزیولوژیکی درگیر در مراقبه را ارائه دادند که کمابیش تنها بر مبنای فعالیت دستگاه عصبی خودکار (ANS) قرار داشت؛ که اگرچه تا اندازهای محدود است، اما نشانگر اهمیت ANS در طی چنین تجربههایی بود (Gellhorn & Kiely, 1972). ایدۀ این نویسندگان بر آن بود که تحریک شدید هر یک از دستگاه سمپاتیک و یا پاراسمپاتیک اگر استمرار یابد، میتواند سرانجام به تخلیه همزمان این دو دستگاه بینجامد (چیزی که میتوان آن را «پیشرفت» دستگاه دیگر به حساب آورد). چند پژوهش غلبۀ فعالیت پاراسمپاتیک در طی مراقبه را نشان دادهاند، که با کاهش میزان ضربان قلب و فشار خون، ضربآهنگ تنفسی و کاهش سوختوساز اکسیژن همپیوند است (Travis, 2001). با اینهمه، مطالعه تازهای از دو شیوه مراقبهای مستقل، با نشان دادن افزایش تغیرپذیری ضربان قلب در طی مراقبه، برانگیختگی دو سویه دستگاههای سمپاتیک و پاراسمپاتیک را پیشنهاد کرد (Peng et al., 1999). فرضیه این پژوهش بر آن بود که افزایش تغییر در ضربان قلب، برانگیختگی هر دو بازوی دستگاه عصبی خودکار را بازتاب میدهد. همچنین این نظر با توصیف ویژه حالات مراقبهای در قالب پدیدههایی که دربردارنده احساس آرامش و سکون شدید، و نیز هوشیاری چشمگیر هستند، جور در میآید. همچنین، نظریه برانگیختگی متقابلِ هر دو بازوی دستگاه عصبی خودکار با پیشرفتهای اخیر در مطالعه برهمکنشهای خودکار سازگار است (Hugdahl, 1996).
فعالیت شبکۀ سروتونی[73]
برانگیختگی دستگاه عصبی خودکار میتواند به تحریک شدید ساختارهایی در هیپوتالاموس جانبی و دسته پیشمغزِ میانی[74] منجر شود، که میدانیم هرگاه مستقیما تحریک میشوند احساسات جذبهای و سرخوشی و بهجت تولید میکنند (Olds & Forbes, 1981). تحریک هیپوتالاموس جانبی همچنین میتواند به تغییراتی در فعالیت شبکۀ سروتونی بینجامد. در واقع، چندین پژوهش نشان دادهاند که پس از مراقبه، فروپاشی فرآوردههای خراب سروتونین (ST) در ادرار بهطرز چشمگیری افزایش مییابد، که نشانگر افزایش کلی ST در طی مراقبه است (Walton et al., 1995). سروتونین یک تعدیلکننده عصبی است که به طور متراکم مراکز بینایی لوب گیجگاهی را تأمین میکند؛ همانجا که سروتونین با قوت تمام بر جریان پیوندهای بینایی تولیدشده توسط این ناحیه اثر میگذارد (Joseph, 1990). سلولهای ستیغ خلفی، ST را هنگامی که توسط هیپوتالاموس جانبی تحریک میشوند، (Rasmussen,1987&Aghajanian, Sprouse) و نیز هنگامی که توسط کرتکس پیشپیشانی فعال میشوند (Juckel, Mendlin & Jacobs, 1999)، تولید و توزیع میکنند. افزایش نسبی سطح ST با عاطفه مثبت همبستگی دارد، در حالیکه سطح پایین ST، اغلب نشاندهنده افسردگی است (Van Praag & De Haan, 1980). این پیوند در خصوص تأثیرات داروهای مهارکنندۀ بازجذب انتخابی سروتونین،[75] که بهطور گستردهای برای درمان افسردگی استفاده میشوند، آشکارا نشان داده شده است. همچنین باید یادآوری کنیم که چند مطالعه بالینی دریافتهاند که اعمال مراقبهای و معنویتمحور میتوانند میزان افسردگی یا بازگشت به افسردگی را بکاهند. پیوند بین اعمال معنوی و کاهش افسردگی، نقش سروتونین در اعمال معنوی را تأیید میکند.
با اینهمه، هنگامی که گیرندههای قشریST (بهویژه در لوبهای گیجگاهی) فعال میشوند، تحریک میتواند منجر به توهمزایی شود. روانگردانهای تریپتامین، همچون سیلوسیبین، و ال اس دی، بهنظر میرسد بیشترین استفاده را از این سازوکار برای تولید تداعیهای بصری شگفتآورشان میبرند (Aghajanian & Marek, 1999). سازوکار چنین رخدادی بهنظر این است که ST جلوی هسته زانویی جانبی را میگیرد، که اطلاعات بصری فراوان قابل عبور را تولید میکند (Funke & Eysel, 1995; Yoshida, Sasa, & Takaori, 1984). ST اگر با بازداری هسته مشبک از زانویی جانبی همراه شود، ممکن است سیالی تداعیهای بصری گیجگاهی را در غیاب درونداد حسی افزایش دهد، که احتمالا به تصویرسازی ذهنی از درون تولیدشدهای منجر میشود که در طی حالات خاص مراقبهای مورد توصیف قرار میگیرند.
افزایش سطح ST میتواند شبکههای عصبشیمیایی دیگر را نیز تحت تأثیر قرار دهد. افزایش سروتونین تأثیری تعدیلگر بر دوپامین دارد، که نشانگر پیوند میان شبکههای سروتونی و دوپامینی است، و میتواند احساسهای سرخوشی و نشئه را افزایش دهد ( Vollenweider, Vontobel, Hell, & Leenders, 1999)؛ احساساتی که فراوان در طی حالات مراقبهای گزارش میشوند. پژوهشها نشان دادهاند که ST بههمراه افزایش گلوتامات، هستههای قاعدهای را به رها کردن استیل کولین برمیانگیزاند، که تأثیر تعدیلگری مهمی در سرتاسر کرتکس دارد (Manfridi, Brambilla, & Mancia, 1999). پژوهشها نشان دادهاند که افزایش استیل کولین در لوبهای پیشانی سامانه توجهی را میافزایند و در لوبهای آهیانهای جهتیابی را بدون تغیر دادن درونداد حسی افزایش میدهند. گرچه هیچ مطالعهای نقش استیل کولین را در مراقبه نکاویده است، اما بهنظر میرسد که این انتقالدهنده عصبی میتواند بخش توجهی، و نیز پاسخ جهتیاب را در برابرِ آورانبرداری تدریجی و پیشرونده درونداد حسی به لوبهای آهیانهای در طی مراقبه افزایش دهد. افزایش ST، در ترکیب با عصبگیری هیپوتالاموس جانبی از غده صنوبری، میتواند این آخری را به افزایش تولید هورمون عصبی ملاتونین (MT) از تبدیل ST بکشاند (Moller, 1992). مشخص شده است که ملاتونین بر دستگاه عصبی مرکزی فشار میآورد و حساسیت به درد را میکاهد (Shaji & Kulkarni, 1998). در طی مراقبه، معلوم شده است که MT پلاسمای خون بهطور ناگهانی افزایش مییابد (Tooley, Armstrong, Norman, & Sali, 2000)، که این میتواند به احساس آرامش و کاهش آگاهی مراقبهکننده از درد کمک کند (Dollins et al., 1993). در شرایط برانگیختگی شدید، آنزیمهای صنوبری نیز میتوانند به شکل درونزاد، توهمزای قدرتمند 5ـمثِکسیـدیمتیلتریپتامین[76] (DMT) را تولید کنند (Monti & Christian, 1981). چندین پژوهش DMT را با انواع گوناگون حالات رازآلود عرفانی، همانند تجربههای خلع بدن، قلب و اعوجاج زمان و مکان، و تعامل با موجودات فوق طبیعی درپیوند دانستهاند (Strassman & Clifford, 1994; Strassman, Clifford, Qualls &Berg, 1996). بنابراین تحریک بیش از حد غده صنوبری در این مرحله میتواند همچنین به تولید DMT بینجامد، که چهبسا با انواع گسترده و متنوع تجربههای عرفانگونی مرتبط باشد که با آن توهمزا درپیوند هستند.
نتیجهگیری: تجربه معنوی در پیشه روانشناختی
با آنکه فصلهای دیگر این کتاب پیوندهای گوناگون معنویت و روانشناسی میکاوند، اما این مقاله نکات بسیار مهمی را پیرامون ماهیت این رابطه پیش میافکند. با نگاه به تاریخ میبینیم که جامعه غربی بر اهمیت علیت، پیشرفتهای تکنولوژیکی، و تجربهگرایی پافشاری ورزیده است. از پایگاه این ارزشها است که پزشکی، روانپزشکی و روانشناسی غربی گسترش یافتهاند. بهنظر ما (با چشمپوشی از دلالت ضمنی مفهوم معنویت در جامعه غربی)، تجربههای عرفانی، مراقبهای، فرآیندهایی طبیعی و چهبسا قابل اندازهگیری هستند که هماره مردمان اقوام ، ادیان و فرهنگهای گوناگون آن را تجربه کردهاند و همچنان نیز تجربه خواهند شد. امکان وجود انواع گوناگونی از ساختارهای نوروساکولوژیکی در بین دارندگان تجربههای معنوی هست. باخبری و حساسیت به باورهای معنوی و فلسفی برای متخصصان بالینی نیز بسیار مهم است (Worthington, McCullough, & Sandage, 1996). متخصصان باید قادر به تمییز نهادن میان رشد معنوی سالم و بهنجار از آسیبشناسی روانی باشند. امیدواریم که تحلیل نوروسایکولوژیکی ترسیمشده در بالا توانسته باشد تمایز بین تجربههای معنوی «بههنجار» و حالات آسیبشناختی را در نظر گرفته باشد. در واقع، این عنوانگذاری میتواند برای تحلیلهای روانشناختی آتی از تجربههای دینی سودمند باشد. با اینهمه، این واقعیت که تجربههای معنوی بر کارکرد [دستگاه عصبی] خودکار، و نیز دیگر فرایندهای شناختی و هیجانی متأثر از قشر مخ اثر مینهد، نشانگر آن است که چنین تجربههایی نه تنها روان آدمی را تحت تأثیر قرار میدهند، بلکه میتوان آنها را استادانه برای کمک به درمان اختلالهای گوناگون نیز بهکار گرفت. پیشتر نشان داده شد که نیایش و مراقبه میتوانند پارامترهای جسمانی و روانشناختی را بهبود بخشند (Carson, 1993; Kabat-Zinn, Lipworth, & Burney, 1985; Kaplan, Goldenberg, & Galvin-Nadeu, 1993; Worthington et al., 1996). همبستههای نوروسایکولوژیکی نهفته در تجربههای معنوی هرچه بیشتر مورد فهم قرار گیرند، این تجربهها را بهتر و بیشتر میتوان تجزیه و تحلیل نمود و در کار بالینی بهکار گرفت. از اینرو تجربه معنوی در طبابت روانشناختی بالینی و روانپزشکی میتواند بسیار پرفایده باشد. همچنین خودِ متخصصان بالینی میتوانند در کمک به بیمارانشان در جهت رشد و تعالی شخصی و معنوی، با بحث از اعمال گوناگون مراقبهای و/یا معنوی، و تشویق آشکار بیماران به توجه و بازگشت به این اعمال، مفید و وسیلهساز باشند. بنا بر نظر روانشناس انسانگرا، روئِن(1983)، این خود[77] است که حلقه گمشده بین [درمان] روانشناختی و معنوی است. نتیجه آنکه، این سخنِ درستی بهنظر میآید که تجربههای معنویای چون مراقبه و دعا و نیایش را میتوان بهعنوان ضمیمهای کمککار به اقدامات درمانیِ غربی افزود. همچنین، رشد معنوی شخص میتواند بخش مهمی از رشد روانیـاجتماعی و نیز نوروسایکولوژیکی وی باشد.
سپاسگذاری: از توماس جی. فیکز[78] برای نکات سودمندش درباره این فصل سپاسگذاریم.
افزوده مترجم: باید توجه داشت، دامی که بر سر اینگونه تحقیقات وجود دارد تقلیلگرایی است؛ بهعبارتی این مطالعات همبستگی را در حد مطالعات علّی ارج نهادن و همبستههای فیزیوژیکیِ تجربههای عرفانی را علت آنها شماردن. بنابراین بهلحاظ روش تحقیق، تحقیقاتی از این دست را صرفاً باید از چارچوب همبستگی نگاه کرد، و نه بیشتر.
.
.
مآخذ
Adair, K. C., Gilmore, R. L., Fennell, E. B., Gold, M.,&Heilman, K. M. (1995). Anosognosia during
intracarotid barbiturate anaesthesia: Unawareness or amnesia for weakness. Neurology, 45,
241–243.
Aghajanian, G. K.,&Marek, G. J. (1999). Serotonin and hallucinogens. Neuropsychopharmacology,
21, 16S–23S.
Aghajanian, G., Sprouse, J.,&Rasmussen, K. (1987). Physiology of the midbrain serotonin system. In
- Meltzer (Ed.), Psychopharmacology: The third generation of progress (pp. 141–149). New
York: Raven Press.
Albin, R., & Greenamyre, J. (1992). Alternative excitotoxic hypotheses. Neurology, 42, 733–738.
Armony, J. L.,&LeDoux, J. E. (2000). How danger is encoded: Toward a systems, cellular, and computational
understanding of cognitive-emotional interactions in fear. In M. S. Gazzaniga (Ed.),
The new cognitive neurosciences (pp. 1073–1074). Cambridge, MA: MIT Press.
Bogen, J. E. (1969). The other side of the brain: II. An appositional mind. Bulletin of Los Angeles Neurological
Society, 34, 135–162.
Bucci, D. J., Conley, M., & Gallagher, M. (1999). Thalamic and basal forebrain cholinergic connections
of the rat posterior parietal cortex. Neuroreport, 10, 941–945.
Bucke, R. M. (1961). Cosmic consciousness. Secaucus, NJ: Citadel Press.
Burton, H., & Jones, E. G. (1976). The posterior thalamic region and its cortical projections in new
world and old world monkeys. Journal of Comparative Neurology, 168, 249–302.
Carson, V. B. (1993). Prayer, meditation, exercise, special diets: Behaviors of the hardy person with
HIV/AIDS. Journal of the Association of Nurses in AIDS Care, 4, 18–28.
Cheramy, A., Romo, R.,&Glowinski, J. (1987). Role of corticostriatal glutamatergic neurons in the
presynaptic control of dopamine release. In M. Sandler, C. Feuerstein,&B. Scatton (Eds.), Neurotransmitter
interactions in the basal ganglia. New York: Raven Press.
Cornwall, J.,&Phillipson, O. T. (1988). Mediodorsal and reticular thalamic nuclei receive collateral
axons from prefrontal cortex and laterodorsal tegmental nucleus in the rat. Neuroscience Letter,
88, 121–126.
Crosson, B., Sadek, J. R., Maron, L., Gokcay, D., Mohr, C. M., Auerbach, E. J., Freeman, A. J., Leonard,
- M.,&Briggs, R.W. (2001). Relative shift in activity from medial to lateral frontal cortex
during internally versus externally guided word generation. Journal of Cognitive Neuroscience,
13, 272–283.
d’Aquili, E. G. (1978). The neurobiological bases of myth and concepts of deity. Zygon, 13, 257–275.
d’Aquili, E. G. (1983). The myth–ritual complex:Abiogenetic structural analysis. Zygon, 18, 247–269.
d’Aquili, E. G. (1986). Myth, ritual, and the archetypal hypothesis: Does the dance generate the word?
Zygon, 21, 141–160.
d’Aquili, E. G., & Newberg, A. B. (1993). Religious and mystical states: A neuropsychological substrate.
Zygon, 28, 177–200.
d’Aquili, E. G.,&Newberg, A. B. (1999). The mystical mind: Probing the biology of religious experience.
Minneapolis, MN: Fortress Press.
Davies, E., Keyon, C. J., & Fraser, R. (1985). The role of calcium ions in the mechanism of ACTH
stimulation of cortisol synthesis. Steroids, 45, 551–560.
Davis, M. (1992). The role of the amygdala in fear and anxiety. Annual Review of Neuroscience, 15,
353–375.
Destexhe, A., Contreras, D.,&Steriade, M. (1998). Mechanisms underlying the synchronizing action of corticothalamic feedback through inhibition of thalamic relay cells. Journal of Neurophysiology,
79, 999–1016.
Dollins, A. B., Lynch, H. J., Wurtman, R. J., Deng, M. H., Kischka, K. U., Gleason, R. E., &
Lieberman, H. R. (1993). Effect of pharmacological daytime doses of melatonin on human
mood and performance. Psychopharmacology, 112, 490–496.
Elias, A. N., Guich, S., & Wilson, A. F. (2000). Ketosis with enhanced GABAergic tone promotes
physiological changes in transcendental meditation. Medical Hypotheses, 54, 660–662.
Fernandez-Duque, D., & Posner,M. I. (2001). Brain imaging of attentional networks in normal and
pathological states. Journal of Clinical Experimental Neuropsychology, 23, 74–93.
Fish, D. R., Gloor, P., Quesney, F. L.,&Olivier, A. (1993). Clinical responses to electrical brain stimulation
of the temporal and frontal lobes in patients with epilepsy. Brain, 116, 397–414.
Foote, S. (1987). Extrathalamic modulation of cortical function. Annual Review of Neuroscience, 10,
67–95.
Frith, C. D., Friston, K., Liddle, P. F., & Frackowiak, R. S. (1991). Willed action and the prefrontal
cortex in man:Astudy with PET. Proceedings of the Royal Society of London, 244, 241–246.
Funke, K., & Eysel, U. T. (1995). Possible enhancement of GABAergic inputs to cat dorsal lateral
geniculate relay cells by serotonin. Neuroreport, 6, 474–476.
Gazzaniga, M. S., & Hillyard, S. A. (1971). Language and speech capacity of the right hemisphere.
Neuropsychologia, 9, 273–280.
Gellhorn, E.,&Kiely,W. F. (1972). Mystical states of consciousness: Neurophysiological and clinical
aspects. Journal of Nervous and Mental Disease, 154, 399–405.
Herzog, H., Lele, V. R., Kuwert, T., Langen, K.-J., Kops, E. R., & Feinendegen, L. E. (1990–1991).
Changed pattern of regional glucose metabolism during yoga meditative relaxation. Neuropsychobiology,
23, 182–187.
Hugdahl, K. (1996). Cognitive influences on human autonomic nervous system function. Current
Opinion in Neurobiology, 6, 252–258.
Infante, J. R., Peran, F., Martinez, M., Roldan, A., Poyatos, R., Ruiz, C., Samaniego, F.,&Garrido, F.
(1998). ACTH and beta-endorphin in transcendental meditation. Physiology and Behavior, 64,
311–315.
Ingvar, D. H. (1994). The will of the brain: Cerebral correlates of willful acts. Journal of Theoretical
Biology, 171, 7–12.
Janal, M., Colt, E., Clark, W., & Glusman, M. (1984). Pain sensitivity, mood and plasma endocrine
levels in man following long-distance running: Effects of naxalone. Pain, 19, 13–25.
Jevning, R.,Wallace, R. K.,&Beidebach, M. (1992). The physiology of meditation:Areview.Awakeful
hypometabolic integrated response. Neuroscience Biobehavioral Review, 16, 415–424.
Jevning, R.,Wilson, A. F.,&Davidson, J. M. (1978). Adrenocortical activity during meditation. Hormones
and Behavior, 10, 54–60.
Jevtovic-Todorovic, V., Wozniak, D. F., Benshoff, N. D., & Olney, J. W. (2001). A comparative
evaluation of the neurotoxic properties of ketamine and nitrous oxide. Brain Research, 895,
264–267.
Joseph, R. (1990). Neuropsychology, neuropsychiatry, and behavioral neurology. New York: Plenum
Press.
Juckel, G. J., Mendlin, A.,&Jacobs, B. L. (1999). Electrical stimulation of rat medial prefrontal cortex
enhances forebrain serotonin output: Implications for electroconvulsive therapy and transcranial
magnetic stimulation in depression. Neuropsychopharmacology, 21, 391–398.
Kabat-Zinn, J., Lipworth, L.,&Burney, R. (1985). The clinical use of mindfulness meditation for the
self-regulation of chronic pain. Journal of Behavioral Medicine, 8, 163–190.
Kaplan, K. H., Goldenberg, D. L., & Galvin-Nadeu, M. (1993). The impact of a meditation-based
stress reduction program on fibromyalgia. General Hospital Psychiatry, 15, 284–289.
Karnath, H. O., Ferber, S.,&Himmelbach, M. (2001). Spatial awareness is a function of the temporal
not the posterior parietal lobe. Nature, 411, 950–953.
Kiss, J., Kocsis, K., Csaki, A., Gorcs, T. J., & Halasz, B. (1997). Metabotropic glutamate receptor in GHRH and beta-endorphin neurons of the hypothalamic arcuate nucleus. Neuroreport, 8,
3703–3707.
Kjaer, T.W., Bertelsen, C., Piccini, P., Brooks, D., Alving, J.,&Lou, H. C. (2002). Increased dopamine
tone during meditation-induced change of consciousness. Cognitive Brain Research, 13(2), 255–
259.
Lazar, S.W., Bush, G., Gollub, R. L., Fricchione, G. L., Khalsa, G.,&Benson, H. (2000). Functional
brain mapping of the relaxation response and meditation. Neuroreport, 11, 1581–1585.
Livesey, J. H., Evans, M. J., Mulligan, R., & Donald, R. A. (2000). Interactions of CRH, AVP and
cortisol in the secretion of ACTH from perifused equine anterior pituitary cells: “Permissive”
roles for cortisol and CRH. Endocrinology Research, 26, 445–463.
Luria, A. R. (1966). Higher cortical functions in man. New York: Basic Books.
Lynch, J. C. (1980). The functional organization of posterior parietal association cortex. Behavioral
Brain Sciences, 3, 485–499.
Manfridi, A., Brambilla, D.,&Mancia, M. (1999). Stimulation of NMDA and AMPA receptors in the
rat nucleus basalis of Meynert affects sleep. American Journal of Physiology, 277, R1488–R1492.
Moller,M. (1992). Fine structure of pinealopetal innervation of the mammalian pineal gland. Microscope
Research Technology, 21, 188–204.
Monti, J. A.,&Christian, S.T. N.-N. (1981). Dimethyltryptamine: An endogenous hallucinogen. International
Review of Neurobiology, 22, 83–110.
Mountcastle, V. B. (1976). The world around us: Neural command functions for selective attention.
Neurosciences Research Progress Bulletin, 14, 1–47.
Mountcastle, V. B., Motter, B. C.,&Andersen, R. A. (1980). Some further observations on the functional
properties of neurons in the parietal lobe of the waking monkey. Brain Behavioral Sciences,
3, 520–529.
Nebes, R. D.,&Sperry,R.W. (1971). Hemispheric disconnection syndrome with cerebral birth injury
in the dominant arm area. Neuropsychologia, 9, 249–259.
Newberg, A. B., Alavi, A., Baime, M., Pourdehnad, M., Santanna, J., & d’Aquili, E. G. (2001). The
measurement of regional cerebral blood flow during the complex cognitive task of meditation: A
preliminary SPECT study. Psychiatry Research: Neuroimaging, 106, 113–122.
Newberg, A. B.,&d’Aquili, E. G. (1994). The near death experience as archetype: A model for “prepared”
neurocognitive processes. Anthropology of Consciousness, 5, 1–15.
Newberg, A. B.,&Iversen, J. (2003). The neural basis of the complex mental task of meditation: Neurotransmitter
and neurochemical considerations. Medical Hypotheses, 61, 282–291.
Newman, J., & Grace, A. A. (1999). Binding across time: The selective gating of frontal and
hippocampal systems modulating working memory and attentional states. Consciousness and
Cognition, 8, 196–212.
O’Halloran, J. P., Jevning, R.,Wilson, A. F., Skowsky, R.,Walsh, R. N.,&Alexander, C. (1985). Hormonal
control in a state of decreased activation: Potentiation of arginine vasopressin secretion.
Physiology and Behavior, 35, 591–595.
Olds, M. E.,&Forbes, J. L. (1981). The central basis of motivation: Intracranial self-stimulation studies.
Annual Review of Psychology, 32, 523–574.
Pardo, J. V., Fox, P. T.,&Raichle, M. E. (1991). Localization of a human system for sustained attention
by positron emission tomography. Nature, 349, 61–64.
Peng, C. K., Mietus, J. E., Liu, Y., Khalsa, G., Douglas, P. S., Benson, H.,&Goldberger, A. L. (1999).
Exaggerated heart rate oscillations during two meditation techniques. International Journal of
Cardiology, 70, 101–107.
Pietrowsky, R., Braun, D., Fehm, H. L., Pauschinger, P.,&Born, J. (1991). Vasopressin and oxytocin
do not influence early sensory processing but affect mood and activation in man. Peptides, 12,
1385–1391.
Poletti, C. E.,&Sujatanond, M. (1980). Evidence for a second hippocampal efferent pathway to hypothalamus
and basal forebrain comparable to fornix system: A unit study in the monkey. Journal
of Neurophysiology, 44, 514–531.
Portas, C. M., Rees, G., Howseman, A. M., Josephs, O., Turner, R.,&Frith, C. D. (1998). A specific
role for the thalamus in mediating the interaction attention and arousal in humans. Journal of
Neuroscience, 18, 8979–8989.
Posner, M. I., & Petersen, S. E. (1990). The attention system of the human brain. Annual Review of
Neuroscience, 13, 25–42.
Pribram, K. H. (1973). The primate frontal cortex: Executive of the brain. In K. H. Pribram & A. R.
Luria (Eds.), Psychophysiology of the frontal lobes. New York: Academic Press.
Redding, F. K. (1967). Modification of sensory cortical evoked potentials by hippocampal stimulation.
Electroencephalography and Clinical Neurophysiology, 22, 74–83.
Renaud, L. P. (1996). CNS pathways mediating cardiovascular regulation of vasopressin. Clinical and
Experimental Pharmacology and Physiology, 23, 157–160.
Rowan, J. (1983). The real self and mystical experiences. Journal of Humanistic Psychology, 23(2), 9–27.
Saver, J. L., & Rabin, J. (1997). The neural substrates of religious experience. Journal of Neuropsychiatry
and Clinical Neuroscience, 9, 498–510.
Shaji, A. V., & Kulkarni, S. K. (1998). Central nervous system depressant activities of melatonin in
rats and mice. Indian Journal of Experimental Biology, 36, 257–263.
Sim, M. K.,&Tsoi,W. F. (1992). The effects of centrally acting drugs on the EEG correlates of meditation.
Biofeedback Self Regulation, 17, 215–220.
Smart, N. (1958). Reasons and faiths: An investigation of religious discourse, Christian and non-
Christian. London: Routledge & Kegan Paul.
Smart, N. (1967). History of mysticism. In P. Edwards (Ed.), Encyclopedia of philosophy. London:
Macmillan.
Smart, N. (1969). The religious experience of mankind. London: Macmillan.
Sperry, R. W., Gazzaniga, M. S., & Bogen, J. E. (1969). Interhemispheric relationships: The
neocortical commisures; syndromes of hemisphere disconnection. In P. J. Vinken&C.W. Bruyn
(Eds.), Handbook of clinical neurology, Vol. 4. Amsterdam: North Holland.
Stace, W. T. (1961). Mysticism and philosophy. London: Macmillan.
Strassman, R. J.,&Clifford, R. (1994). Dose–response study of N,N-Dimethyltrypamine in humans:
- Neuroendocrine, autonomic, and cardiovascular effects. Archives of General Psychiatry, 51,
85–97.
Strassman, R. J., Clifford, R., Qualls, R., & Berg, L. (1996). Differential tolerance to biological and
subjective effects of four closely spaced doses of N,N-Dimethyltrypamine in humans. Biological
Psychiatry, 39, 784–795.
Sudsuang, R., Chentanez, V.,&Veluvan, K. (1991). Effects of Buddhist meditation on serum cortisol
and total protein levels, blood pressure, pulse rate, lung volume and reaction time. Physiology
and Behavior, 50, 543–548.
Thomas, A. G., Vornov, J. J., Olkowski, J. L., Merion, A. T.,&Slusher, B. S. (2000). N-Acetylated alpha-
linked acidic dipeptidase converts N-acetylaspartylglutamate from a neuroprotectant to a
neurotoxin. Journal of Pharmacology and Experimental Therapies, 295, 16–22.
Tooley, G. A., Armstrong, S. M., Norman, T. R., & Sali, A. (2000). Acute increases in night-time
plasma melatonin levels following a period of meditation. Biological Psychology, 53, 69–78.
Travis, F. (2001). Autonomic and EEG patterns distinguish transcending from other experiences during
transcendental meditation practice. International Journal of Psychophysiology, 42, 1–9.
Turner, V. (1969). The ritual process: Structure and anti-structure. Ithaca, NY: Cornell University
Press.
Van Bockstaele, E. J.,&Aston-Jones, G. (1995). Integration in the ventral medulla and coordination
of sympathetic, pain and arousal functions. Clinical and Experimental Hypertension, 17, 153–
165.
Van Praag, H., & De Haan, S. (1980). Depression vulnerability and 5–Hydroxytryptophan prophylaxis.
Psychiatry Research, 3, 75–83.
Vogt, B. A., Finch, D. M., & Olson, C. R. (1992). Functional heterogeneity in cingulate cortex: The
anterior executive and posterior evaluative regions. Cerebral Cortex, 2, 435–443.
Vollenweider, F. X., Leenders, K. L., Scharfetter, C., Antonini, A., Maguire, P., Missimer, J.,&Angst,
- (1999). Metabolic hyperfrontality and psychopathology in the ketamine model of psychosis
using positron emission tomography (PET) and [18F]fluorodeoxyglucose (FDG). European
Neuropsychopharmacology, 7, 9–24.
Vollenweider, F. X., Vontobel, P., Hell, D.,&Leenders, K. L. (1999). 5-HT modulation of dopamine
release in basal ganglia in psilocybin-induced psychosis in man—a PET study with [11C]raclopride.
Neuropsychopharmacology, 20, 424–433.
Walton, K. G., Pugh, N. D., Gelderloos, P.,&Macrae, P. (1995). Stress reduction and preventing hypertension:
Preliminary support for a psychoneuroendocrine mechanism. Journal of Alternative
Complementary Medicine, 1, 263–283.
Waterhouse, B. D., Moises, H. C.,&Woodward, D. J. (1998). Phasic activation of the locus coeruleus
enhances responses of primary sensory cortical neurons to peripheral receptive field stimulation.
Brain Research, 790, 33–44.
Weingartner, H., Gold, P., Ballenger, J. C., Smallberg, S. A., Summers, R., Rubinow,D. R., Post, R. M.,
& Goodwin, F. K. (1981). Effects of vasopressin on human memory functions. Science, 211,
601–603.
Worthington, E. L., McCullough, M. E.,&Sandage, S. J. (1996). Empirical research on religion and
psychotherapeutic processes and outcomes: A 10-year review and research prospectus. Psychological
Bulletin, 119, 448–487.
Yadid, G., Zangen, A., Herzberg, U., Nakash, R.,&Sagen, J. (2000). Alterations in endogenous brain
beta-endorphin release by adrenal medullary transplants in the spinal cord. Neuropsychopharmacology,
23, 709–716.
Yoshida, M., Sasa, M., & Takaori, S. (1984). Serotonin-mediated inhibition from dorsal raphe neurons
nucleus of neurons in dorsal lateral geniculate and thalamic reticular nuclei. Brain Resolution,
290, 95–105.
Ziegler, D. R., Cass,W. A.,&Herman, J. P. (1999). Excitatory influence of the locus coeruleus in hypothalamic–
pituitary–adrenocortical axis responses to stress. Journal of Neuroendocrinology, 11,
361–369.
[1] Andrew B. Newberg, Stephanie K. Newberg؛ این مقاله برگرفته از این کتاب است:
Handbook of the psychology of religion and spirituality / edited by Raymond F. Paloutzian, Crystal L. Park. 2005 The Guilford Press.
[2] biomedical؛ بخشی از پزشکی که با علوم طبیعی، بهویژه زیستشناسی، زیست شیمی، و زیست فیزیک سروکار دارد. م
[3] neurophysiological؛ عصبْ روانشناسی یا روانشناسی عصبنگر. م
[4] Homo sapiens؛ انسان هوشمند. م
[5] religiogenic
[6] neuropsychological؛ نوروسایکولوژی، به تبیین رابطه مغز و رفتار میپردازد و میخواهد به چگونگی کنش مغز پیببرد، بهطور مثال چه سازوکارهایی در تفکر، یادگیری و احساس اهمیت دارند و چگونه راهاندازی میشوند و چه تأثیری بر رفتار انسان دارند. این شاخه علمی، مطالعات خود را بیشتر روی بیماران مبتلا به اختلالهای مغزی متمرکز میکند و از تغییر رفتار چنین بیمارانی به اهمیت ساختها و روابط آنها در بروز رفتاری معین پی میبرد. نوروسایکولوژی به دو زیرمجموعه بالینی و تجربی طبقهبندی میشود. اولی بر بیماران و دومی چگونگی کنش مغز افراد سالم را بررسی میکند. برای آشنایی مقدماتی با این شاخه جذاب این دو کتاب بسیار رهگشا هستند: مقدمات نوروسایکولوژی از دکتر داوود معظمی؛ نوروسایکولوژی و سایکوفیزیولوژی از دکتر محمد کریم خداپناهی، از دو از انتشارات سمت. م
[7] neurobiology؛ یا زیستشناسی اعصاب، که عبارت است از مطالعه سلسله اعصاب و نقش آن در رفتار. م
[8] cognitive operator
[9] cognitive modules
[10] explanatory models
[11] neuroanatomical؛ نوروآناتومی یا تشریح اعصاب، مطالعه ساختمان سلسلهاعصاب، اجزای تشکیلدهنده آن، و ارتباطات آنها است. م
[12] این گمانهورزی مولفان به تمامه نوعی فلسفهورزی است و از شآن علم خارج است و نیازمند مباحث دقیق فلسفی است که آیا میتوان از آن دادههای علمی این نتیجه فلسفی را گرفت. بهنظر میرسد که چنین نباشد. م
[13] holistic operator
[14] gestalt
[15] larger contextual framework
[16] parietal lobe in the right (or nondominant) hemisphere
[17] neuroanatomical substrate
[18] more reductionist processes
[19] group ritual
[20] individual contemplation؛ گرچه، استفاده از برابرنهادِ تأمل/ژرفاندیشی در برابر contemplation رایج است؛ اما به نظر میرسد که رسا نباشد. شاید در متون مربوط به معنویت «در خود رَوی» معادل بهتری باشد، که ناشی از خودکاوی برای معرفتالنفس و در نتیجه یافتن خدا و معادلهای آن درفرهنگهای گوناگون است. م
[21] community؛ (معادل از آقای داریوش آشوری). نمونه واضح این حالات را در شعر زیر از بین عربی میبینیم: لقد صار قلبی قابلاً کلَ صورۀ/ فمرعی لغزلان و دیر لرهبان/ و بیت لأوثان و کعبۀ طایف/ و الواح توراۀ و مصحف قرآن/ اَدین بدینِ الحُب انی توجهت/ رکایبه فالحب دینی و ایمانی. حلّاج نیز میگوید: نیک در ادیان اندیشه کردم و دریافتم که آنها یک اصلِ واحدند با شاخههای فراوان: تفکّرت فیالادیان جدّ تحقق/ فألفیتها اُصلاً شعباً جمّا. م
[22] Absolute Unitary Being
[23] “average” person
[24] structural aspects of a society
[25] communitas tribus
[26] myth-ritual experience
[27] اشاره به آدرس یادشده در کمی پیشتر: (AUB; d’Aquili & Newberg, 1999).
[28] neurochemical.
[29] posterior superior parietal lobe
[30] Neurotransmitters؛ نوروترنسمیتر/ عصبرسانه. م
[31] Activation of the Prefrontal and Cingulate Cortex
[32] prefrontal cortex
[33] cingulate gyrus
[34] positron-emission tomography ؛ نقشهبرداری شیمیایی و هستهای از مغز و نخاع بر اساس متابولیسم مواد در سلولها. م
[35] innervation؛ عصبگیری، توزیع عصبی و عصبرسانی به یک عضو، غده یا عضله. م
[36] lateral geniculate and lateral posterior nuclei
[37] striate cortex
[38] posterior superior parietal lobule
[39] photon emission computed tomography
[40] deafferentation؛ وارونه و برگرفته از afferent بهمعنای آوران، که به هدایت تکانههای عصبی از محیط (اعضای حسی) به سوی سلسله اعصاب مرکزی اطلاق میشود. راههای آوران به آن دسته از راههای عصبی گفته میشود که اطلاعات را از گیرندهها به سلسله اعصاب مرکزی میرسانند. در مقابل efferent قرار دارد از واژهلاتین بهمعنای دورکننده که در نوروفیزیولوژی به هدایت تکانههای عصبی از سلسله اعصاب مرکزی به محیط (عضلات، غدد) اطلاق میشود. نورونهای وابران اطلاعات را به اندامهای مجری هدایت میکنند و غالباً نورونها یا راههای محرکه نامیده میشود. م
[41] dopaminergic؛ یا: دوپامینرژیک. مشخصکننده یا مربوط به راهها، الیاف یا نورونهایی که در آنها دوپامین یک ناقل عصبی است؛ شبکه فعالیت دوپامین. م
[42] basal ganglia؛ تودههایی از ماده خاکستری که در عمق هز یک از نیمکرهها مدفون هستند که خود از اجسام مخطط و کپسول داخلی تشکیل میشوند. م
[43] glutamatergic؛ یا: گلوتاماترژیک. مشخصکننده یا مربوط به راهها، الیاف یا نورونهایی که در آنها گلوتامات یک ناقل عصلی است. م
[44] Yoga Nidra meditation
[45] GABAergic؛ یا گابارژیک. مشخصکننده یا مربوط به راهها، الیاف یا نورونهایی که در آنها اسید گاما آمینوبوتریک، یک ناقل عصبی است. م
[46] Balint’s syndrome؛ نوعی آگنوزی بینایی که در آن بیمار قادر به تشخیص تصاوری منفرد هست، اما وقتی یک سری تصویر به او نشان داده شود، از تشخیص توالی آنها ناتوان است. م
[47] perception
[48] neocortical؛ نئوکورتکس، تازهترین قسمت قشر مخ پستانداران (از نظر تکاملی) است که شامل پالئوکورتکس و آرکیکوکورتکس که لایههای سلولس معدودتری نسبت یه شش لایه نئوکورتکس دارند، نمیشود. م
[49] nucleus accumbens
[50] functional magnetic resonance imaging
[51] paragigantocellular nucleus of the medulla
[52] locus coeruleus؛ ناحیهکوچک متمایل به رنگ آبی در تگمنتوم خلفیـجانبی پل دماغی، که با چشم غیرمسلح نیز قابل رؤیت است. م
[53] cellular “noise”
[54] cerebral decrease
[55] paraventricular، تحریک این هسته با نمک یا الکتریسیته موجبه میل به آشامیدن میشود.
[56] adrenocorticotropic
[57] caudal ventral medulla
[58] GABAergic inhibition
[59] vasoconstrictor arginine vasopressin
[60] endorphin؛ این اصطلاح مخفف اندروفینهای درونزا است. اندورفینها مواد شبه مرفین هستند که در مغز تولید میگردند. این مواد نفش مهمی در کنترل رفتارهای هیجانی، نظیرحالات مربوط به درد، اضطراب، تنش، ترس و حالات عاطفی وابسته که از درد ناشی میشوند دارند. م
[61] هیپوتالاموس 11 هسته دارد که در هستههای قدامی، میانی و خلفی تقسیمبندی میشوند. م
[62] naloxone؛ یک آنتاگونیست اوپیومی که موجب برگشت آثار فارماکولوژیک مورفین و داروهای شبیه مورفین با رقابت بر سر محلهای گیرنده در مغز میگردد. م
[63] N-methyl-D-aspartate receptors
[64] excitotoxic processes؛ یک فرایند آسیبشناختی که توسط آن سلولهای عصبی با تحریک بیش از حد توسط نوروترنسمیترهایی از قبیل گلوتامات آسیب دیده و یا کشته میشوند.
[65] N-acetylated alpha-linked acidic dipeptidase
[66] endogenous NMDAr antagonist N-acetylaspartylglutamate
[67] ketamine؛ یک داروی هوشبر با تأثیر سریع و کوتاه که خصوصیات روانگرا شبیه فن سیکلیدین دارد که آن نیز یک هوشبر و ضد درد است که سابقا در اعمال جراحی استفاده میشد. م
[68] nitrous oxide؛ داروی هوشبر و مخدر ملایمی که به گار خندهآور مشهور است. م
[69] out-of-body
[70] Autonomic Nervous System؛ سیستم عصبی خودمختار نیز گفته میشود. م
[71] Gellhorn
[72] Kiel
[73] Serotonergic Activity؛ یا: سروتونرژیک. م
[74] median forebrain bundle
[75] selective serotonin reuptake inhibitor medications؛ دارووهایی که سروتونین را انتخاب می کنند و مانع بازجذب آنها میشوند. م
[76] 5-methoxy-dimethyltryptamine
[77] self
[78] Thomas G. Fikes
.
.
عصبروانشناسی (نوروسایکولوژی) تجربه دينی و معنوی
نویسندگان: اندرو بی. نیوبرگ ، استفانی کی. نیوبرگ
ترجمه محسن زندی (پژوهشگر روانشناسی دین)
.
.